Mouse anti double-stranded RNA (K2)
Catalog number: 10030010/10030005
| Clone | K2 |
| Isotype | IgM kappa |
| Product Type | Monoclonal Antibody |
| Units | 10 ml/5 ml |
| Host | Mouse |
| Application | dsRNA-immunoblotting ELISA Immunofluorescence Sandwich ELISA |
Background
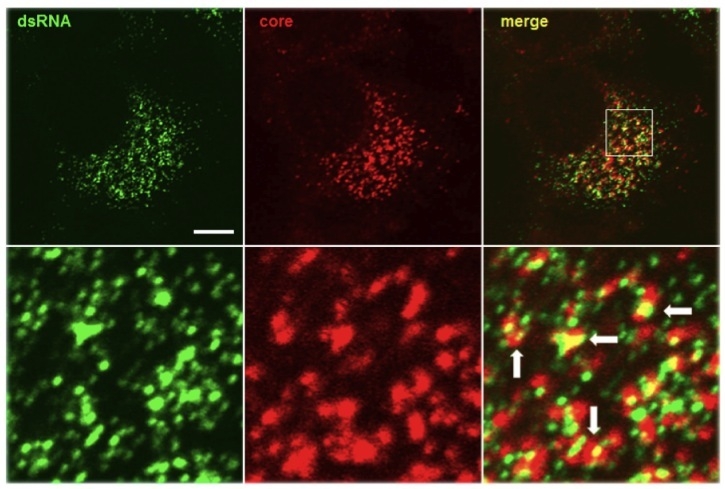
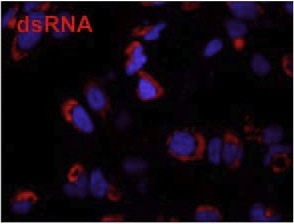
Over the past decade our double-stranded RNA (dsRNA)antibodies have been used extensively to detect and characterise plant and animal viruses with dsRNA genomes or intermediates. In addition, the anti-dsRNA antibodies can be used as a diagnostic tool to detect pathogens, including detection in paraffin-embedded fixed tissue samples (Richardson et al. 2010). K2 anti-dsRNA monoclonal antibody has an IgM isotype. K2 has primarily been used in ELISA and sandwich ELISA. Generally, K2 can be used in applications where an anti-dsRNA antibody with an isotype other than IgG (IgG2a) is required.
Synonyms: Mouse anti dsRNA
Source
Female DBA/2 mice were injected intraperitonially with a mixture of 50 ug L-dsRNA and 75 ug methylated bovine serum albumin, emulsified in complete Freund's adjuvant. After several boosts spleen cells were fused with Sp2/0-Agl4 myeloma cells to generate the hybridoma clone.
Product
The mAb K2 recognises double-stranded RNA (dsRNA) provided that the length of the helix is greater than or equal to 40 bp dsRNA. Recognition is independent of the sequence and nucleotide composition of the antigen. All naturally occurring dsRNAs investigated up to now (4050 species) as well as poly(I).poly(C) and poly(A).poly(U) have been recognised by K2. As described by Schönborn et al. K2 binds with high avidity to all dsRNAs investigated.
Formulation: Culture supernatant (RPMI, 5% fetal calf serum).
Concentration: Undiluted hybridoma supernatant
Applications
MAb K2 is primarily used for a sandwich ELISA to detect and quantitate (after calibration) dsRNA (see Schönborn et al.). For this application it should be diluted with PBS. It may also be advantageous to use K2 for immunofluorescence studies. The optimum working dilution of the antibody for any specific application should be established by titration. Not for use for clinical purposes. For in vitro use only.
Storage
After delivery antibodies should be aliquoted and stored at -20 ° or -70 °C. After adding 10 mM sodium azide undiluted antibody can also be stored at +4 °C for a short period of time. For long term storage the mAb should be kept frozen. Repeated freezing/thawing cycles should be avoided.
Shipping Conditions: The IgM mononclonal antibody K2 is only sold as hybridoma supernatant and shipped at ambient temperature.
Caution
This product is intended FOR RESEARCH USE ONLY, and FOR TESTS IN VITRO, not for use in diagnostic or therapeutic procedures involving humans or animals. It may contain hazardous ingredients. Please refer to the Safety Data Sheets (SDS) for additional information and proper handling procedures. Dispose product remainders according to local regulations.This datasheet is as accurate as reasonably achievable, but Exalpha Biologicals accepts no liability for any inaccuracies or omissions in this information.
References
1) Schönborn, J., Oberstrass, J., Breyel, E., Tittgen, J., Schumacher, J. and Lukacs, N. (1991) Monoclonal antibodies to double-stranded RNA as probes of RNA structure in crude nucleic acid extracts. Nucleic Acids Res.19, 2993-3000. 2) Lukacs, N. (1994) Detection of virus infection in plants and differentiation between coexisting viruses by monoclonal antibodies to double-stranded RNA. J. Virol. Methods 47, 255-272. 3) Lukacs, N. (1997) Detection of sense:antisense duplexes by structurespecific anti-RNA antibodies. In: Antisense Technology. A Practical Approach, C. Lichtenstein and W. Nellen (eds), pp. 281-295. IRL Press, Oxford. 4) S. J. Richardson, A. Willcox, D. A. Hilton, S. Tauriainen, H. Hyoty, A. J. Bone, A. K. Foulis, N. G. Morgan. Use of antisera directed against dsRNA to detect viral infections in formalin-fixed paraffin-embedded tissue. J Clin Virol. (2010) 49(3); 180-5. doi: 10.1016/j.jcv.2010.07.015.